

Abstract
The gravity force results in the human structure having weight. The skeleton supports this, but speculation regarding tensegrity as the human structural strategy brings up new questions. This article attempts to develop this conceptualization and offers a framework of thought in order to better understand how humans hold their shape and move about. A working model is built using both the research to date as well as the general application of the principles of tensegrity and hydrostatics to human architecture. An attempt is made to explain why and how the skeleton, as a rule, is not intended to sit on itself. The possibility that many degenerative symptoms, including joint wear, are actually the result of structural system failure is considered.This article was previously printed in the June 2012 issue of Structural Integration published by the Rolf Institute®.
The Elephant in the Living Room Fascia is the organ of posture. Nobody ever says this; all the talk is about muscles. Yet this is a very important concept, and because this is so important, we as Structural Integrators must understand both the anatomy and physiology, but especially the anatomy of fascia. !e body is a web of fascia . . . For example, why, when we work with the superficial fascia does this change the tone of the fascia as a whole? Ida P. Rolf, Ph.D. (Feitus, 1978, p. 124) Structural integration (SI) practitioners are constantly confronted with the medical evaluative and treatment histories of their clients and sometimes, things just don’t seem to stack up. For instance, a review of the literature on the biomechanics of bone reveals that although many biomechanical properties have been measured (e.g., compressive, tensional, shearing, bending, and torsional strengths), the skeleton is primarily investigated as a defense against gravitational force. Also, the mechanical properties of bone as a material and a tissue have been intensely explored and reported on, but many of the conclusions admit that these results are only suggestive of conditions in vivo. Individual bones are complex aggregates of tissues and properties and are variable along their length and breadth. Whole skeletons, as groupings of different bones, are therefore more difficult to make generalizations of. This so confounds scientific evaluation in vivo that biomechanical studies of bones have been mostly done on individual bones or pieces of mineralized bony material. How forces are transmitted through and across the human structure remains something of a frontier. This discussion admittedly faces the same obstacles, but the system as a whole can and needs to be conceptualized functionally.
Compression, tensile resistance, bending resistance, torsional strain, and stresses on bone have all been investigated. Absent are abrasion or wear resistance studies on bone although cartilage has been analyzed this way again and again. THis author has found no evidence of inquiry into why cartilage wears out, although it might be assumed that these studies have been done because cartilage wear is common. Cartilage is understood to be miraculously smooth with a frictional coefficient so small that it has yet to be successfully mimicked with prosthetics. This is the only protection for joint surfaces, and for many people lasts a lifetime. In others, it wears prematurely. It would seem that regardless of how smooth the cartilaginous surface and how effective a lubricant is synovial fluid, constant rubbing over a lifetime would wear it out. One must wonder why all hips and knees don’t wear out. Is it simple genetics? In those who require replacements, how come bilateral replacements are not always necessary? The evolutionary path of prosthetic joints as extremely hard, abrasion-resistant appliances points to the expectation of high compression and frictional loads. Assuming that abrasive wear is normal, this is a reasonable response to the presentation of many orthopedic patients. Obviously, titanium is far more resistant to wear than mineralized bone. Why, then, are titanium prosthetics expected to last on the order of only ten years while bones are designed for a lifetime of effective use? It is a matter of logic to see that in a normal situation, joint surfaces must not rub against each other; ideally, space exists between opposing cartilaginous surfaces, but these must approximate with unusual load (jumping, for instance). Perhaps the need for a prosthetic is not normal, but indicative of a structural system failure whose most prominent measureable symptom is joint wear. Once the joint is replaced without attending to the system failure, the load and abrasive wear continues unchecked—or is even increased as a consequence of the replacement, hence the need for materials like titanium.
The Tensegrity Component
The exploration of tensegrity as the architectural strategy for life has been increasing. It has been shown to be the case on a cellular level (Ingber, 1997) and has been speculated to be operative on an organism level as well (Langevin, 2006; Ingber, 2008). By definition, the stiffer elements in a tensegrity structure are in approximation, but not in direct contact. The real question is: How does this work in a living body? In other words, if human structure is indeed based on tensegrity, then how do the bones stay apart? We know that the application of the tensegrity model to cells has resulted in a much more precise and satisfying understanding of the architecture of life, thanks in large part to the work of Donald Ingber. We also know that in nature, the smallest design forms are generally repeated in larger format (fractal patterning), so it is a natural progressionto see that human beings are also designed on this basic structural principle. Ths notion, basic to structural integration, has been catching on in the larger scientific community (Chen & Ingber, 1999; Pfluger, 2008). Classically, in SI circles, the debate has been between tensegrity and hydrostatic models for human structure. As time goes on and more research is published on the biomechanical basis for movement and support in life forms, the apparent conclusion is that both are operative. The tensegrity model requires a pre-stressed, continuously tensile, discontinuously compressive structure. It is simple to imagine the bones of the skeleton as representing the stiffer elements on the classic tensegrity design and the connective tissue (CT) as the cable-like elements. Muscles are enclosed within the cabling in order to provide for dynamic motion in a short time frame (Cassuto, 2009).
From an inquiring perspective, there are some big, lingering questions. Tensegrity masts and spoked wheels are all very nice as design models, but it becomes necessary to understand how to make the leap to human form in a way that empowers us to work with precise intent. Wheel hubs maintain a consistent distance from the rim when the spokes are balanced, but applying that to human architecture is not satisfying. Something is missing. Making this harder to grasp, or even accept, is the reality that when muscles contract, they impose mostly compressive forces across these joint spans until motion begins. As the joint changes position, the proportion of compressive force generally diminishes, but that it is a powerful force at initiation is impossible to ignore. Even those considering tensegrity as a whole skeletal system design assume the joints are in direct compressive contact (Chen & Ingber). One would think this doesn’t fit with the system. (NB: many tetrapods, e.g., felines and ungulates, have no bony attachments connecting the axial skeleton to the shoulder girdle, yet these limbs accept and utilize movement loads quite well). Besides, remember that cartilage is too soft a substance to withstand a lifetime of compressive contact, even lubricated by synovial fluid. It is known to have a tremendous capacity to withstand hydrostatic pressures, but seems not to have any adaptive response to compressive forces (Eckstein, Hudelmaier, & Putz, 2006). This article suggests that the joints are generally floating in delicate approximation. They are indeed lubricated, and for that lubrication to be effective, light, non-compressive contact must be the case. Joint capsules help to maintain the higher hydrostatic pressures necessary for this arrangement. The nature of tensegrity design principles would have these stiff elements held in approximation in precise place by the balance of the whole system. Our understanding of the human strategy of tensegrity needs to address this in order to be taken seriously and for our work to progress.
The Hydrostatic Component
The molt stage of crustaceans (e.g., crabs) is a good example of hydrostatics. During the molt, or growth stage, crustaceans lose their hard skeleton, so have to briefly rely on hydrostatic pressure for motion.
Triggered by hormones, the molt-stage crabs pump themselves full of fluid to increase their size (to allow for growing room), their shell opens, and they climb out. Very vulnerable in this stage, they hide until their cuticle hardens to become a new, larger shell. It had been believed that crabs were like rag dolls at this stage, unable to move at all, since their hard exoskeletons were gone. However, more recently it has been shown that during their molt phase crabs are indeed capable of moving about, but their reduced speed and agility renders them more vulnerable to predation (Taylor & Kier, 2003).
Besides, if they were really like rag dolls, how would they have climbed out of their shells in the first place? Their hydrostatic skeleton ‘works’ without the hard parts, but not well enough for their lifestyle. Defending themselves with this softer cuticle would be ineffective at best. In fact, the soft-shell stage cuticle has been shown to have the same tensile strength as the hardened shell, so is able to resist hydrostatic forces in motion (Taylor, Hebrank, & Kier, 2007). The cuticle, resistant to tension, acts as a hydrostatically powered skeleton, enabling movement while it hardens. The basic hydrostatic model works (very well in caterpillars, for example), but skeletal struts give animals more options. Crabs can move more quickly and efficiently with less energy with a hard exoskeleton than they can during their hydrostatic molting phase.
Putting It Together
Looking at things from a hydrostatic point of view, it is simple to see cells as bags of fluid. Less simple to imagine, but equally true, is that whole human beings are bags of viscous fluid as well. If we didn’t have compression-resistant struts, then when we jumped from high places, we’d flatten (as with Ingber’s cells) like pancakes, springing back to shape when the forces were relieved. In fact, when we jump, we do deform a little and return to “zero” when the forces are relieved, but our shapes remain remarkably the same. This is the genius of the system.
Bone is known to be almost equally resistant to compressive and tensile forces (Currey, 1984). If we add hydrostatic pressure to the tensegrity model of struts held in place by the balanced, dynamic tension of the fascial system, there is a very good argument for the maintenance of relative position of the bones to the rest of the system, no matter how we move and how we change our shape and load profile. Perhaps the network of all fascial structures defines boundary containers, from joint capsules to superficial fascia, which acting together provide for whole-structure stability and balanced suspension in dynamic motion. This web provides for the extensive use of hydrostatic pressure, arguably a requirement for living tensegrity structures.
Superficial fascia as a Superficial Connector
Superficial fascia (SF), seen classically in structural integration as a body stocking, covers our bodies, but how exactly is it connected everywhere otherwise?
The answer to this goes in several directions by necessity. Imagine the external layer of fascia, the SF, and deep, or investing fascia (DF) as a double-walled tube. The outer layer is an areolar, more elastic layer. Connected to it by extensible fibers is a deeper, stiffer layer that dives even deeper, or invests, in strategic places.
The fiber direction of the deep fascia runs at right angles to those of the muscles, so is clearly set up to resist expansion. In a cross section of this model, the bones appear suspended within the tube (Figure 1).
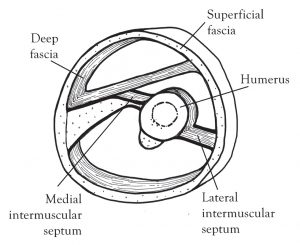
Figure 1. Stylized section through the upper third of right arm, showing connective tissues. Drawings by the author.
Occasionally, these diving layers, or septa, connect to the bones of the skeleton, surround the bones as periosteum, and/or maintain space between parallel bones as an interosseous membrane. This constitutes a completely continuous network, both communicating and distributing load (Benjamin, 2008). The fascial compartments so defined are packed in place primarily with muscle tissue. The high water content of living tissues results in an internal fluid pressure which presses against the septa, generating tensile force everywhere and adding great stability to the entire design (Figure 2).

Figure 2 Stylized section through uppon third right arm as in figure 1, also including muscle tissue and major vessels.
Frei Otto suggested the hydrostatic nature of fascial compartments long ago (Horvitz, 1981). By imagining the structural system this way, it is clearly obvious that muscular contraction communicates across and through all the septa and that any muscular contraction increases the internal pressure locally, further stabilizing the position of the CT tensegrity elements. Huijing and Baan (2008) have proven such coordination of muscular contractions many times. The pre-stress in the SF combined with the hydrostatic nature of our mostly fluid tissues keeps our entire structural system, including its individual compartments, whole and under pressure. This internal pressure maintains reflexive tensile load in the SF, so the system is necessarily reciprocal. It is not possible to overstate the importance of this quality. This is not a new idea. When cutting skin or CT (as in surgery), the tissues immediately give way due to the tension stored within (Guimberteau, Sentucq-Rigall, Panconi, Boileau, Mouton, & Bakhach, 2005). This tension is global.
Muscles = Motion Engines + Packing
When thinking of muscles, one is accustomed to imagining the origin, insertion, and muscle belly of a particular muscle as an individual gross motor unit. Here, attempt to see muscles in cross-section acting not individually, but in groups, part of a compete network of mechanical balance. Imagining them this way, they become not only motors for motion, but generators of the internal fluid pressures necessary to both drive and stabilize body position and movement. Muscles, anatomically not CT themselves, but units whose very fibers are invested with CT, are surrounded by envelopes of fascia. These, in turn are in close association with adjacent envelopes and septa. Having proper tensional balance and resiliency in each fascial envelope allows the motor/muscle unit held within it to function optimally and to produce motion with no lag time. As an individual muscle contracts, its fascial container, or “envelope,” expands and therefore its tendons approximate. This results in an action that both provides for and regulates motion about a joint. What is less obvious but equally vital is the action of muscles on their adjacent fascial compartments (including other myofascial units) and CT structures to influence the precise tensioning of intermuscular septa. It is clear that the expansion of muscle compartments yields precise increases in internal pressure and tensile loading of CT (the neurological implications of this will be considered later). The surrounding SF contains and is strategically joined to the deeper structure at specific places, creating a whole-body balanced, tensional environment.
Physical Implications
The bones, suspended within fascial compartments, are held in place by sub-compartments, which are themselves casings defined by septa and packed with muscle (please refer back to Figure 2). The same pressures resulting from muscular contractions between these septa that cause venous blood to return to the heart also stabilize bony and CT position within structural “tubes,” and by extension, the whole body. Given this conceptualization of bones, it follows that a failure in balance will result in structural symptoms. Bodies have an exceptional capacity to self-regulate, by adjusting tension as necessary, and even in extraordinary circumstances, to distort certain bones (Cassuto, 2009). If even this adaptive threshold is crossed, however, what follows must be a systemwide failure of the tensegrity network.
Thanks to the gravity force, tensegrity degenerates toward compression. In the human body, as tensional balance is lost, the spaces so brilliantly maintained between articular surfaces are diminished enough to wear cartilage and then the bony joint surfaces proper. Seeing things this way, it seems far better to intervene early and restore balance than to allow irreparable degeneration to progress, ultimately requiring joint replacement. If the nature of structural balance in life were more generally recognized and restored when necessary, there might be fewer such traumatic interventions required and perhaps, when joints did needed replacing, they might only need to be replaced once.
Neurological Implications
By regulating internal pressures between deeper fascial tissues, our tensional CT balance makes precise regulation of our position in space possible at any moment and in any “shape.” It has long been established (Stilwell, 1957; Ralston III, et al, 1960; McCredie, 2007) that CT, including bone, is innervated and that proprioception is at least one of its functions. If fascia is seen as a passive tissue, the proprioceptive nervous system function within it can’t be recognized for the powerful tool it is. Rather, there has been speculation as to why these nerves are involved at all. Assuming human structure is tensegrity/hydrostatics based and, therefore, in system-wide constant communication, it is easy to imagine these neurological elements as being proprioceptive in nature and maybe even the basic nature of our proprioceptive system. Loading of any type, from any direction, stimulates these nerve endings and delivers precise information, which in communication with the rest of these tissues tells us where we are in space. An organism-wide tensional balance must be communicative, if even to simply remain in balance. How else would such self-regulation otherwise be possible? CT is a perfect medium for this. As well stated by Helene Langevin (2006, p. 1075-6), “. . . the connective tissue matrix is a key participant in mechanotransduction, or mechanisms allowing cells to perceive and interpret mechanical forces . . . one can envisage the whole body web of connective tissue involved in a dynamic, body-wide pattern of cellular activity fluctuating over seconds to minutes reflecting all externally and internally generated mechanical forces acting upon the body.” Currently, there is great interest in the nature and function of nervous system communication within the CT. The conclusions of work done exploring myofascial force transmission (Huijing & Baan, 2008; Huijing, 2009) also support this possibility. What a perfectly obvious way to determine exactly where we are in space.
Classically, in SI circles, the debate has been between tensegrity and hydrostatic models for human structure. As time goes on . . . the apparent conclusion is that both are operative.
Interventional Implications
Structural therapists look for and attempt to achieve something often referred to as continuity or flow. Colloquially, this is not a conscious physics or engineering notion, although the preceding discussion proves that it clearly physically exists. The therapist’s senses are educated over time to perceive this quality in his/her clients. It is sought after through intentional intervention in the tissues and the whole living system. When contacting the fascial system with a precise touch, but also with a deep, bony intent, this tensional framework allows the facilitation of major positional shifts that are of very deep origin.
This is accomplished with even a vague conceptual understanding, but by using educated, informed touch while knowing the anatomical relationships of the tissues in detail, a practitioner becomes able to determine which fiber groups in which vectors are strained and can use these attachments fluently. The physical and neurological capability of CT, mated with the properties of tensegrity and our structural system’s pull toward equilibration, provides overwhelming potential for therapeutic intervention. Precise, informed touch lights up not only mechanoreceptors, but also neuroreceptors, and is communicative system-wide. When working with clients, the quality of informed presence is ultimately the most powerful tool a practitioner has.
References
Benjamin, M. (2008). The fascia of the limbs and back – a review. Journal of Anatomy, 214(1), 1-18.
Cassuto, S. (2009). A fresh look at tensegrity, or . . . I didn’t grow up playing with a tensegrity mast, did you?
in M. Beech, P. Kemper, & K. Schumaker (Eds.), IASI 2009 yearbook of structural integration. Missoula, MT:
International Association of Structural Integrators, 38-44.
Cassuto, S. (2009). Intra-innominate distortions: making a case, in M. Beech, P. Kemper, & K. Schumaker (Eds.),
IASI 2009 yearbook of structural integration. Missoula, MT: International Association of Structural Integrators, 34-37.
Chen, C. S., & Ingber, D. E. (1999). Tensegrity and mechanoregulation: from skeleton to cytoskeleton. Osteoarthritis and Cartilage, 7(1), 81-94.
Currey, J. (1984). Comparative mechanical properties and histology of bone. American Zoologist, 24(1), 5-12.
Eckstein, F., Hudelmaier, M. & Putz, R. (2006). The effects of exercise on human articular cartilage. Journal of Anatomy, 208(4), 491-512.
Feitus, R. (editor). (1978). Ida Rolf talks about Rolfing and physical reality. Boulder, CO: Rolf Institute.
Guimberteau, J. C., Sentucq-Rigall, J., Panconi, B., Boileau, R., Mouton, P., & Bakhach, J. (2005). Introduction to the knowledge of subcutaneous sliding system in humans. Annales de Chirurgie Plastique Esthetique, 50(1), 19-34.
Horvitz, G. (1981). Pneumatic and tensile structures: the work of Frei Otto. Bulletin of Structural Integration, 7(2), 5-8.
Huijing, P., & Baan, G. (2008). Myofascial force transmission via extra-muscular pathways occurs between antagonistic muscles. Cells Tissues Organs, 188(4), 400-414.
Huijing, P. (2009). Epimuscular myofascial force transmission: a historical review and implications for new research. International Society of Biomechanics Muybridge Award Lecture, Taipei, 2007, published in Journal of Biomechanics, 42, 9-21.
Ingber, D. E. (1997). Tensegrity: the architectural basis of cellular mechanotransduction. Annual Review of Physiology, 59, 575-599.
Ingber, D. E. (2008). Tensegrity and mechanotransduction. Journal of Bodywork and Movement !erapies, 12(3), 198-200.
Langevin, H. (2006). Connective tissue: a bodywide signaling network? Medical Hypotheses, 66(6), 1074-1077.
McCredie, J. (2007). Nerves in bone: the silent partner. Skeletal Radiology, 36(6), 473-475.
Pfluger, C. (2008). The meaning of tensegrity principles for osteopathic medicine. (Master’s thesis for MS in osteopathy submitted at the Donau University, Krems and the Vienna School of Osteopathy.) Retrieved from http://www.osteopathicresearch.com/paper_pdf/ Pflueger.pdf
Ralston III, H. J., Miller, M. R., & Kasahara, M. (1960). Nerve endings in human fasciae, tendons, ligaments, periosteum, and joint synovial membrane. Anatomical Record, 136, 137-147.
Stilwell, D. L. (1957). The innervation of tendons and aponeuroses. American Journal of Anatomy, 100(3), 289-317.
Taylor, J. R., Hebrank J., & Kier W. M. (2007). Mechanical properties of the rigid and hydrostatic skeletons of molting blue crabs, Callinectes sapidus Rathbun. !e Journal of Experimental Biology, 210, 4272-4278.
Taylor, J. R. & Kier, W. M. (2003). Switching skeletons: hydrostatic support in molting crabs. Science, 301(5630),
209-210.Thoughts on Tensegrity and Hydrostatics in Human Architecture[:]